(This post is written to highlight a recent experiment which is now hosted as a preprint in PeerJ Preprints. You can find that paper here. Check it out and provide some open peer review!)
We live in an era of widespread human-driven extinction: the Anthropocene. Its a fact that many more species are being lost now than at any point in recorded history, and the future is grim. We are forecasted to lose 6,300% more species by 2100 than we have lost in the last 66 million years (based on evidence from the fossil record). So naturally there has been intense interest in cataloging the world’s biodiversity, and conducting experiments to understand the consequences of losing all these species. But are we looking at the right metric of diversity?
Species richness has a long love affair with ecologists, going back a century or more. Its simple, interpretable, and behaves well under most statistical scenarios. But it is also glosses over important differences, which, in the right circumstances, makes it incredibly misleading.
Here’s an example. Consider two rocky shore communities, 1 (left) and 2 (right):

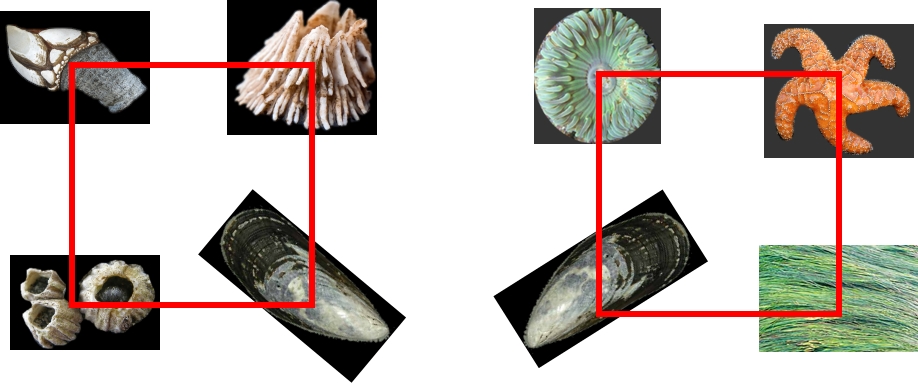
Say our goal is to compare the diversity of these two communities. For community 1, we lay down our quadrat, and count four species:

The gooseneck barnacle (Pollicipes), the thatched barnacle (Semibalanus) and the acorn barnacle (Balanus), as well as the mussel (Mytilus).
For community 2, we also find four species:

The same mussel as above, but also a seastar (Pisaster), an anemone (Anthopleura), and a seagrass (Phyllospadix).
As biologists, we are forced to conclude that the communities have equal diversity. 4 = 4. But intuitively we know something is wrong with this picture: the second community just seems more diverse, doesn’t it? Why is that?
Well, for starters, there are more kinds of things. The first community contains three species of barnacles. While taxonomically distinct (for which read: they are different species!), they are all still barnacles: sessile, shelled, filter feeders. No offense to the humble barnacle, but there is only so many different approaches one can take to being a barnacle, compared the amazing variety of life we see around us everyday.
The second community has the sessile mussel and anemone, but unlike the mussel, the anemone is soft-bodied. The seastar is mobile, and predatory, not to mention much bigger than the other organisms. And the seagrass is not even an animal, its a plant! So what gives?
It all goes back to the implicit assumption we make when we calculate species richness, which is that all species are equally distinct. This means we consider the acorn barnacle to be as different from the thatched barnacle as it is to the seagrass.
This assumption goes against every intuition: clearly the two barnacles are more similar than either is to the seagrass, but we are constrained by our metric. Enter the functional trait.
Despite a long and varied history of interpretation (partly due to varying applications across argonomy and ecology), most ecologists are now in agreement that a functional trait is any measurable aspect of an organism that reflects what it is doing, and how it interacts with other organisms and its environment. These includes aspects of its morphology (e.g., body size), ecology (e.g., diet), environmental tolerances, behavior, and even phenology.
The application of functional traits allows us to score species along a continuum of dissimilarity. Some species share functional traits (they are the same size, eat the same thing, etc.) and thus are more similar than others that do not. Now, if we apply functional traits to the above example, we might find the dissimilarity among species looks more like this for the two communities:
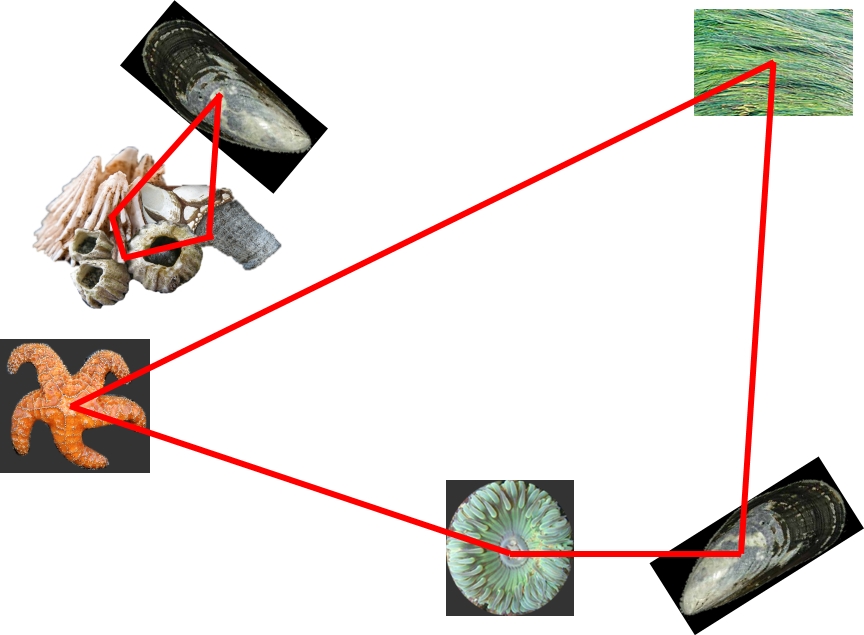 Here, the barnacles are all very similar to one another, and the organisms in the second community are all widely different. We have drawn on our knowledge of the organisms themselves to describe the ‘diversity’ of each community.
Here, the barnacles are all very similar to one another, and the organisms in the second community are all widely different. We have drawn on our knowledge of the organisms themselves to describe the ‘diversity’ of each community.
There is a growing recognition of the benefits of a functional trait approach. Not only does it better discriminate among species in terms of what role they are playing in the community, but also highlights potential for redundancy.
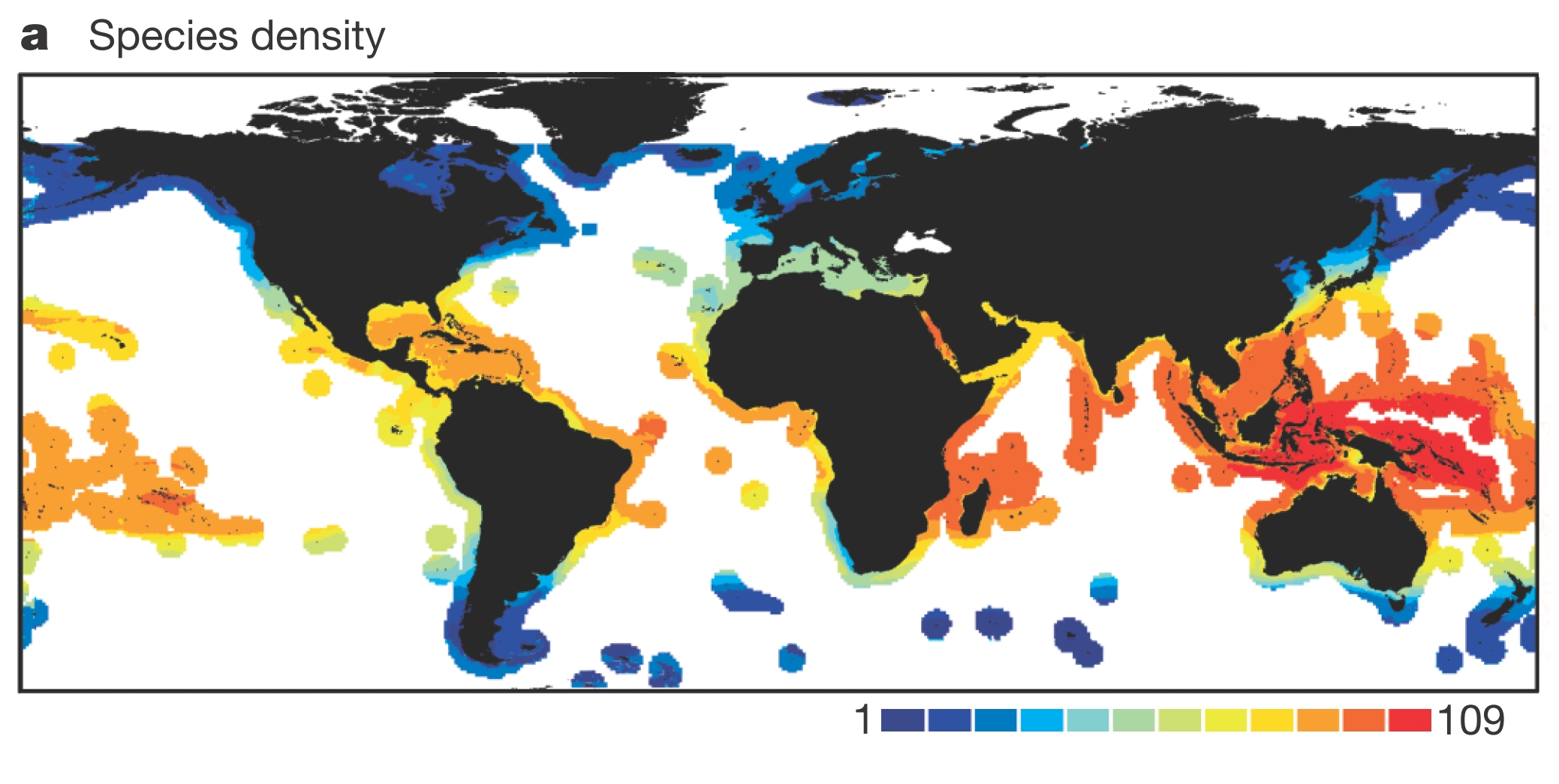
Take an example from our recent publication on global patterns in reef fish diversity. A traditional, species-based approach yields a long known pattern in macroecology: highest diversity at the equator, and decreasing towards the poles, with highest diversity concentrated in the so-called ‘Coral Triangle.’
But if we look at not just richness, but what species are doing, we find a very different pattern: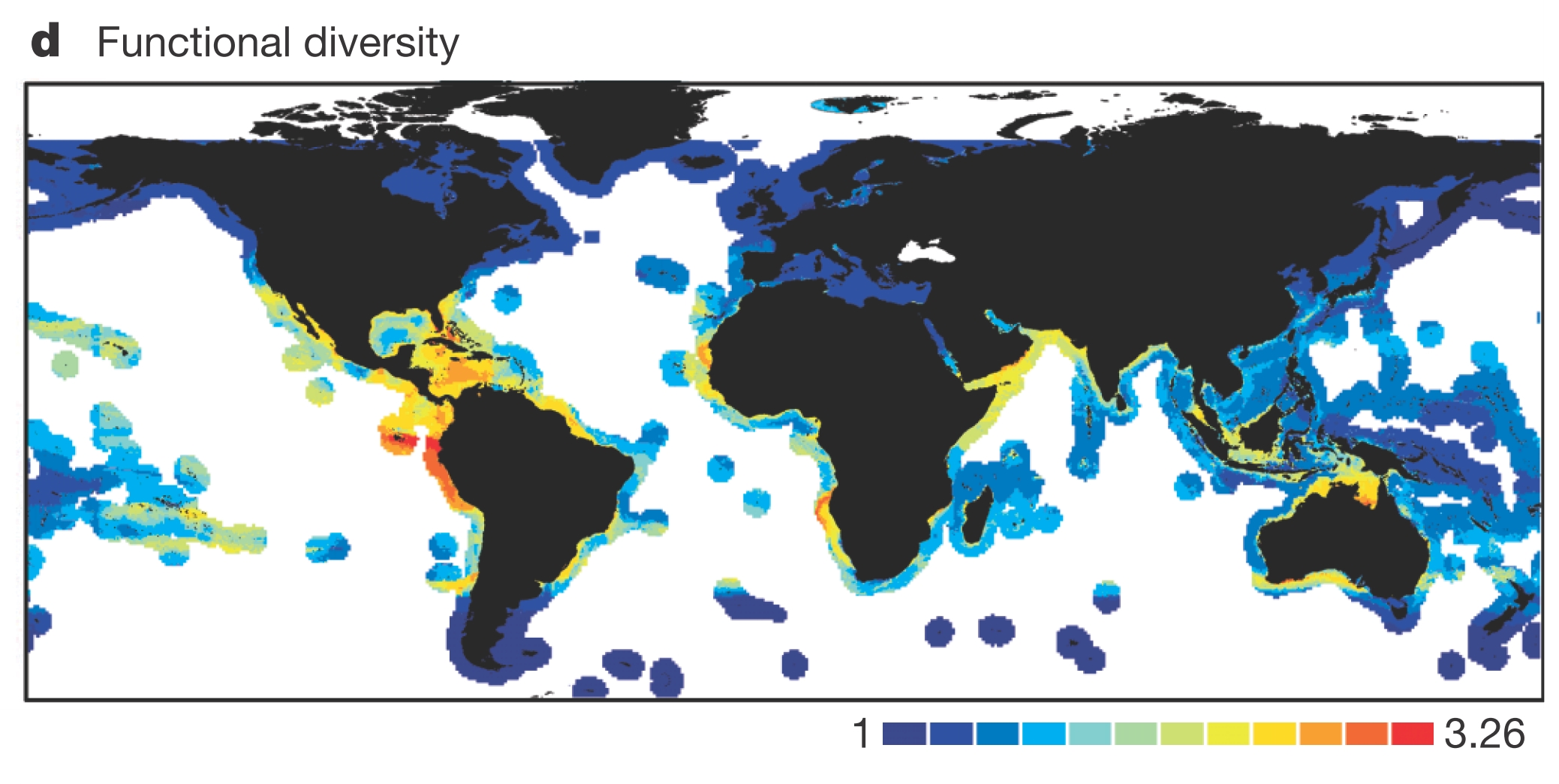
This map suggests that the Coral Triangle is one of the least functionally diverse places on the planet. In other words, it harbors a lot of species, but in general, they are all doing more or less the same thing. (True we know that in an Eltonian world, there must be differences among species in how they acquire and compete for resources that allows them to persist, but we are talking broad strokes here.)
In contrast, hotspots shift towards less rich but functionally diverse areas such as the Galapagos and the southern coast of Australia. This is interesting for a few reasons:
(1) It emphasizes that these area contain unique types of fishes
(2) It emphasizes areas of low redundancy, where every species is doing something very different
Why do we care? Well, we like the idea of seeing crazy fishes (reason #1). But we also recognize that species perform certain roles in the ecosystem: they grow, eat, and sometimes even change their environment. Those roles can be hugely beneficial to humanity by providing food, clean water, and myriad other services that underpin our very society.
If an area is highly speciose, but all of those species are doing the same thing, then we can stand to lose a few species and not lose any functionality. Oppositely, it an area has few species, but all of them are doing something different, then we can’t lose any species without sacrificing some functionality (reason #2). Alternately, we many only have certain functions in regions where certain species appear! For example, we lose a lot erosion control and shoreline buffering when we move out of a salt marsh, eponymously named because of the foundational marsh grasses that compose it.
This is all well and good in principle, but the only way to truly test if this is the case–whether more functionally diverse communities sustain ecosystem functions–is by experimentation: manipulate functional traits and redundancy within levels of richness, and see if it makes a difference for ecosystem functions related to the services above, such as production, consumption, nutrient cycling, and so on.
An added wrinkle to this idea is that the global analysis above incorporated any kind of fish as long as it could be observed. This means that the analysis necessarily spanned many trophic levels. There are likely to be interactions within (e.g., competition) and between (e.g., predation) trophic levels that will affect the ultimate level of functioning. So experiments manipulating functional traits need to understand how functional traits interact both among and between trophic levels to really make inferences above the whole communities we see in nature.
Unfortunately, we are sorely lacking this food web perspective in current functional diversity manipulations, at least until now. In the summer of 2012, I simultaneously manipulated multiple functional traits of crustacean and molluscan herbivores and their fish and crab predators in experimental mesocosms to understand how the community diversity (herbivores and predators together) and the diversity of each trophic level contributed to ecosystem functioning. I recently made this work available as preprint, so please take a look and provide feedback!
Using a model comparison and structural equation modeling approach, I showed that community functional diversity better predicted ecosystem functioning across three trophic levels than species richness alone. This was because there is tremendous functional variation within a certain level of species richness (think the two rocky shore assemblages in the example above).
Since a picture speaks a thousand words, here are two replicate mesocosms, each containing 6 species. On the left is the ‘low’ functional diversity treatment. Notice how scuzzy it is. Now look at the ‘high’ functional diversity treatment on the right. Clean, clear (you can see the bottom!) and a little bit of the algal habitat I placed in at the beginning of the experiment.

Moreover, when I decomposed community functional diversity into separate indicators of herbivore traits and predator traits, two important results emerged.
(1) The functional diversity within a trophic level enhanced the biomass of that trophic level, and
(2) The functional diversity of the predators more effectively depressed the biomass of the herbivores, but only by modifying the functional diversity of the herbivores.
This is really cool, as it reinforces trends from decades of biodiversity-ecosystem function manipulations of species richness, but extends it to functional traits. Also, it implies that multitrophic functional diversity matters to ecosystem functioning.
So what does this mean in practice? It means when we survey two reefs composed of grazers and scrapers and small predators and apex predators, and we find that one community is equally rich but contains a greater diversity of functional traits than another, we can infer enhanced functioning in that reef versus the other based on experimental evidence.
Where do we go from here? I manipulated functional diversity of two trophic levels and nine species. A lot of the functional variation came from whether something was a predator or an herbivore. But there was also evidence that there was subtle variation among very similar grazers. Going after the functional values that delimit those differences is a key frontier.
Also, nine species is a lot relative to most diversity-function experiments, whose median number of species included is only three. But there is so much greater scope for manipulation as the number of species increases, not to mention those experimental assemblages begin to resemble more and more communities found in nature. So increasing the number of species — and perhaps traits — manipulated is a necessity for understanding the role of functional diversity in both depauperate and extremely speciose ecosystems, like those highlighted in the maps above.
Functional diversity is a relatively nuanced way of comparing organisms based on their ecological differences, but its not a panacea. It may depend strongly on which traits are considered, interactions among multiple traits, and what the goals of the analysis are (and whether they may change). Careful thought needs to be put into the choice and use of functional traits, but when they are used appropriately, they have great potential to reveal more about the natural world than ever before.

This is a really great post and the buckets of water are certainly effective visuals!!!
Quite helpful